
plant stem
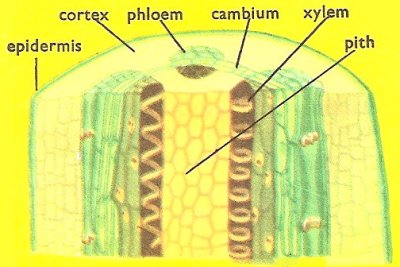
Figure 1. Cross-section of a mature plant stem.
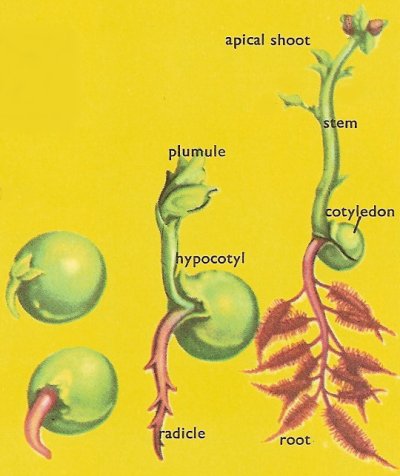
Figure 2. Stages in the germination and early growth of a pea.
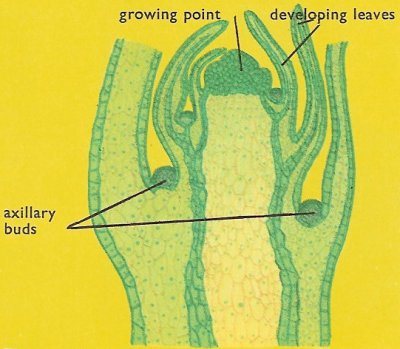
Figure 3. Section of the tip of a stem.
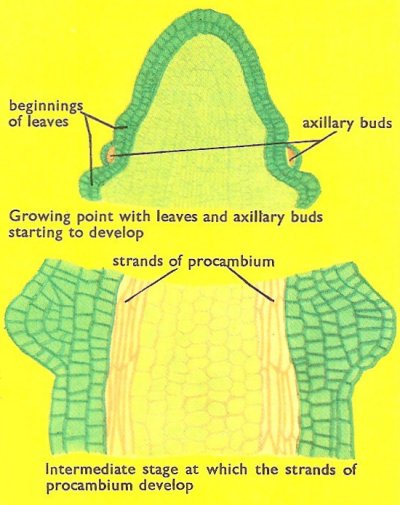
Figure 4. Passage of a stem from embryonic structure to mature structure with differentiated structures.
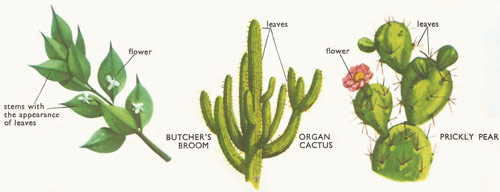
Figure 5. Stems of butcher's broom, organ, and prickly pear.
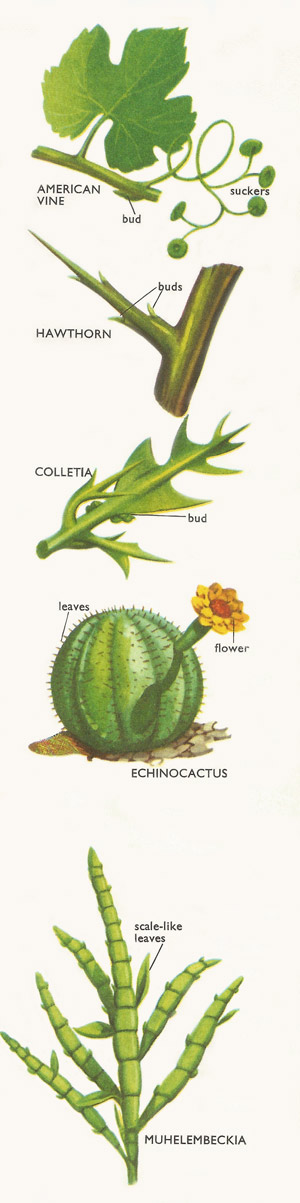
Figure 6. Types of stems.
Plant stems perform two basic functions: they support the leaves and flowers and they carry water and food from place to place within the plant. A typical stem is cylindrical and may be soft (herbaceous) or woody. It is usually branched and leafy. The point at which a leaf joins the stem is called the node. There are several nodes on a stem, each separated by leafless internodes. Each node has one or more leaves, each of which has a bud in its basal angle (axil). These axillary buds often remain dormant but may produce a branch, especially if the main stem is damaged. Buds are in fact miniature shoots (stems) which have not yet elongated. The leaves are clustered together around the tip (the growing point). It is in the winter that buds are most noticeable. The outer leaves are modified as protective scales around the young stem. When these scale-leaves fall off in spring they leave rings of scars on the stem. The age of a woody twig can be established by counting these scar patches.
The main axis of a plant may continue to grow throughout life, giving off branches laterally. This is the racemose branching pattern, well illustrated in larches and "Christmas trees." The cymode branching pattern differs in that the main stem apex does not continue through life. It ends in a flower or dies away and a lateral bud assumes the main position each year.
Internal structure of a stem
The internal structure of the stem reflects the function (Figure 1). There are conducting and supporting tissues. These are basically the same in all parts of the plant and the description given here will cover roots and leaves as well. It is in the arrangement of the tissues that roots and stems mainly differ.
The internal structure is studied by cutting very thin sections and examining them under a microscope. A suitable, easily obtained dicotyledon stem for study is that of the sunflower. A section of an internode cut a short distance from the tip shows the mature primary structure. The epidermal cells are regular and have a waxy cuticle on the outside. The cortex cells are simple and of the type known as parenchyma. In the outer cortical region the cells have thicker walls for extra support and are known as collenchyma cells. The inner layer of the cortex is the starch sheath whose cells have starch deposits within them. Inside this sheath is the stele, made up of vascular tissue and pith. The conducting (vascular) tissue is arranged in vascular bundles. Each bundle has on the outside a number of sclerenchyma fibers. These are elongated cells whose walls have been impregnated with various chemicals known collectively as lignin. The fibers have no cytoplasm and are therefore not living cells. They are tough and elastic, enabling the plant to stand bending by the wind. The phloem is the tissue through which manufactured food is transported. It consists of elongated sieve-tubes with large vacuoles. The ends of the tubes are perforated (sieve-plates), allowing passage of the food solutions. Each sieve tube has a companion cell alongside it which regulates its functioning. There is usually some ordinary parenchyma tissue too.
The dicotyledon bundles are said to be open (i.e., they contain cambium). Cambium is the name given to the actively dividing cells or meristems in the center of the bundle. These play an important part in secondary growth. The inner part of the bundle consists of the water-conducting tissue called xylem. These are elongated vessels and shorter tracheids, although the latter are not common. Xylem tissues are dead and have lignified walls. They are not evenly thickened however. The thickness is spiral or ringed in the first-formed xylem (protoxylem). The later vessels have numerous unthickened pits through which the water passes to neighboring vessels. The pith and the medullary rays are normally of simple parenchyma cells.
Monocotyledon stems differ considerably from the foregoing. The bundles are not arranged in any regular pattern and are of the closed type, i.e., there is no cambium. Strengthening tissue may occur, especially near the outside. The bundles are frequently surrounded by sclerenchyma fibers.
Plant growth
When a seed planted in the soil germinates, the embryo that is formed is elongate in shape. At the tip of the part which grows upwards there is an apical shoot, whose growth increases the height of the stem. The downward-growing tip will penetrate the soil and branch to form a network of roots. In both cases, actual growth takes place just behind the tip not along the whole length of the stem and root (Figure 2).
The growing point is surrounded and protected by the developing leaves. In the axils of the leaves axillary buds develop into new growing points. The growing point in a stem is situated right at the tip. It consists of rapidly dividing meristematic cells which are without vacuoles (Figure 3).
Growth takes place just behind the tip. The cells produced by the meristem develop vacuoles and lengthen rapidly, producing an elongation of the stem. As the cells lengthen they begin to differentiate, or change in character. The outer cells develop into epidermal cells and very quickly tiny leaves arise and cover the growing point. Other cells give rise to the cortex, while certain columns of cells retain their ability to divide and are known as pro-cambium strands (Figure 4).
In the elongating region the first conducting tissues (protoxylem and protophloem) are formed from the procambial strands. The vascular tissues lengthen very rapidly to form the conducting strands. At about the same time the first strengthening tissues (sclerenchyma fibers) develop on the outside of the phloem. The procambial strand continue to produce vascular tissue beyond the elongating region until only a narrow band of cambium remains.
Secondary thickening occurs to some degree in most dicotyledons and is especially marked in those with perennial (persistent) aerial parts (i.e. woody plants). It involves the laying down of extra vascular and strengthening tissue necessitated by the increased size of the plant. Some of the cells of the medullary rays become active again and join with the cambium of the vascular bundles to form a meristematic ring. Secondary xylem and phloem are then formed. The xylem tissues build up inside the cambium and force it and the phloem further from the center. A tree trunk consists mainly of xylem tissue, with a thin layer of phloem near the outside. In areas with well-marked seasons the xylem shows annual rings. In the spring, when the sap is rising, the xylem consists mainly of large vessels, but in autumn there is a high proportion of fibers.
In cross section spring and and autumn wood are very different. producing the ringed effect. Secondary phloem does not build up a thick layer because the cells are unthickened and are sooner or later crushed by the pressure of the expanding xylem tissue. This tension usually breaks down the epidermis too and its protective function is taken over by the phellum. This is a corky tissue formed from a layer of cortex which becomes meristematic. The phellum is a dead tissue and cuts off food from all tissues outside it. These tissues die and as a whole they form the bark of a tree.
Modified stems
Stems frequently perform other functions in addition to or instead of their basic ones. The only thing to remember about a stem is that it is any part of the plant from which leaves, flowers, or buds grow, so that they can be very diverse indeed and may often quite strange (Figures 5 and 6).
The chief modifications are concerned with food storage and vegetative reproduction. Runners are long thin stems which grow over the ground surface and produce new plants some distance from the parent. Strawberries reproduce rapidly in this way. Horizontally-growing underground stems are termed rhizomes. They may be food stores, as in irises, or merely reproductive shoots, as in some grasses and weeds, such as couch-grass and bindweed.
The 'eyes' of a potato are buds, so the potato is also a stem. Potatoes are swellings (tubers) arising on the underground parts of the stem. They serve as food storage and reproductive organs. These underground stems can be distinguished from roots by the presence of scale leaves and buds.
Corms are special types of underground stems. They are swollen with food reserves and carry a number of scale leaves. One or more buds are developed in the axils of the leaves and these buds produce the flowering shoot. The base of each new shoot forms a new corm. Corms are distinct from bulbs. The latter are small underground stems with a number of fleshy scales in which food material is stored. In both corms and bulbs the flowering shoot is fully formed underground and grows rapidly when environmental conditions are suitable. Next year's shoot develops in the axil of one of this year's leaves.
Climbing stems are fairly common. They may climb by coiling round a support (e.g., runner beans), by using tendrils (e.g., vines), or by using hooked prickles to scramble over the vegetation (e.g., blackberry). Protective spines often occur as stem modifications (hawthorn) or as stem outgrowths (rose). Stems are in some cases modified to perform as leaves. Butcher's broom has very leaf-like branch stems and its leaves are tiny scales. The stems of cacti also act as leaves (and water stores too). Stems acting as leaves are called phylloclades.
Strange stems
A great variety of plant structures which grow above the ground must be regarded as stems, although they do not look like them. The illustrations below show a few of the more striking and curious of these; most strange stems have grown in these shapes for a definite reason, such as adapting the plant to local conditions.
Stems with suckers
In the American vine (Vitis inconstans) some shoots are transformed into tendrils with flattened suckers at the ends. These have the power of sticking to any surface they meet. The suckers enable the plant to cling to trees or rocks and so to climb up into the light and air.
Spine-like stems
If we examine a hawthorn bush or an anchor plant (Colletia) we find it has many sharp thorns. These have tiny scale-like leaves, which show that they are really stems. The thorns are a good protection against browsing herbivorous (plant-eating) animals.
Round or spherical stems
In some plants of the cactus family, such as Echinocactus and mammillaria, the whole plant is spiny and shaped like a ball. Here the spines are leaves whose shape has altered and the 'ball' is a stem. This inflated stem serves to store water in the very dry deserts where these cactus grow. The leaves are reduced to thorns to prevent loss of water by evaporation and, of course, to give protection too.
Ribbon-shaped stems
In the curious plant called Muhelembeckia the stem has the shape of a long ribbon, from which sprout a few scale-like leaves. This is another example of stems being altered in structure in order to reduce loss of water.
Spatulate and columnar stems
The flattened, bat-shaped stems of the prickly pear (Opuntia) and the thick, column-like stems of the organ cactus (Cereus) are both designed to store water, as these are desert plants. The leaves are reduced to spines as in other cacti.
Leaf-like stems
Anyone looking at a plant of the butcher's broom (Ruscus aculeatus) would suppose that it has the normal arrangement of leaves growing out of a stalk. But if you look closely at the 'leaves' you will see that little white flowers grow out of the centers of them. They cannot therefore be leaves - they are stems transformed to look like leaves and to perform the function of both stems and leaves.
Where is the chlorophyll?
We know that plants make their food and live by means of a process called photosynthesis that is carried out with the help of the green substance chlorophyll in their leaves. In plants in which the leaves are lost or reduced to spines the stem is always green and takes over this essential function from the leaves.

