
bivalve
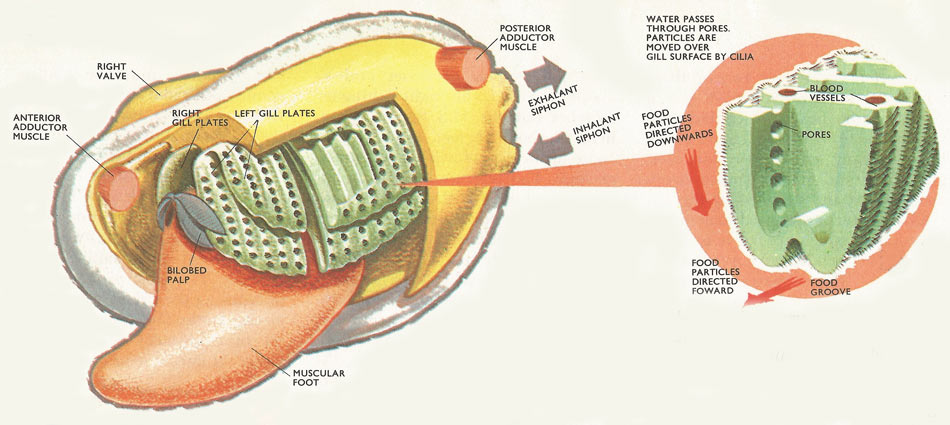
Figure 1. The left valve and mantle lobe of a bivalve mollusk has been removed leaving the gill plates and the palps exposed. A section of the gill plate has been cut away, showing the structure of the gills more clearly. Insert: detailed section of the gills showing cilia, pores, and food grooves.
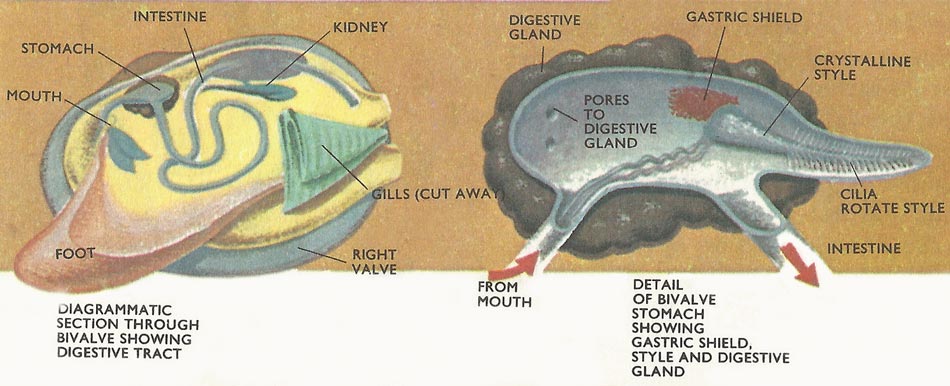
Figure 2. Mollusk digestive tract.
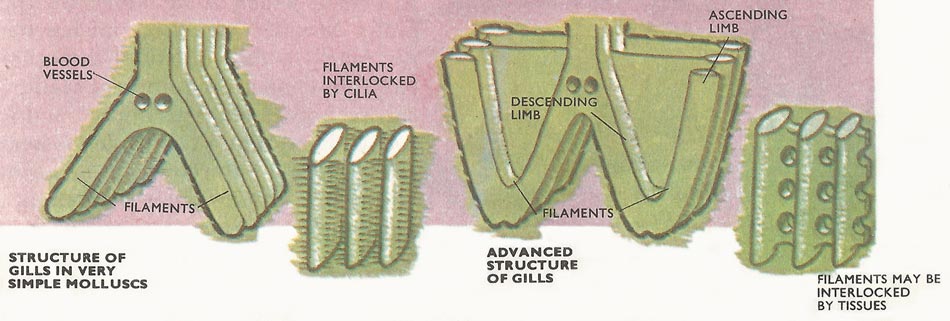
Figure 3. In the simplest molluscs – the protobranches – the gills remain simple. The 'stalk' gives off filaments on each side and these join to form two plates. In more advanced bivalves, each filament grows downwards and then upwards so that a descending and ascending section is formed. Two plates on each side are formed. Water for respiration moves inside through pores but food particles are retained on the surface by cilia.

Figure 4. Giant clam (Tridacna gigas) – the largest and most massive of all bivalves. It is found in coral reefs in tropical seas. A single valve (one side of the shell) may weigh as much as 200 pounds.
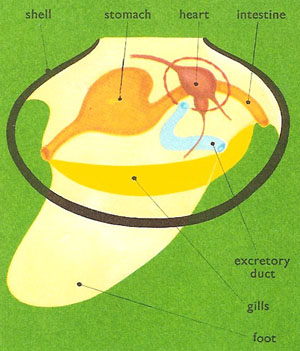
Figure 5. Anatomy of a bivalve.
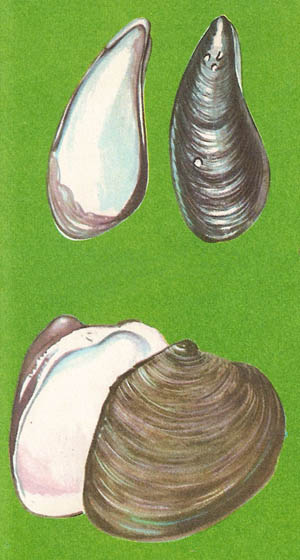
Figure 6. (Top) Mussel (Mytilus edulis), a bivalve that anchors itself to rocks by a bunch of threads called abyssus. (Bottom) Venus shell (Venus mercenaria), a bivalve that buries in mud and breathes through a pair of fleshy tubes call siphons. Water enters through one tube and is expelled through the other.
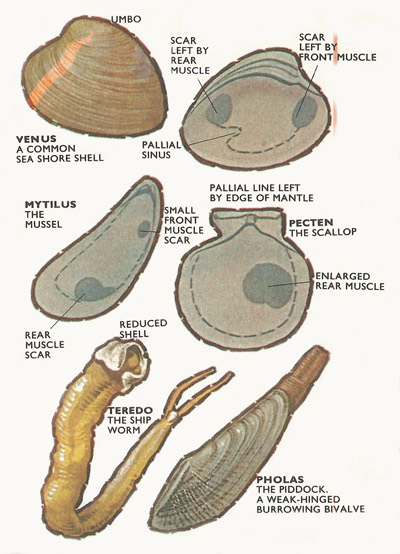
Figure 7. Top: Shell of Venus. Where the muscles abut against the shell, scars are left. The pallial line marks the edge of the mantle lobe. An insertion of the pallial lone marks the position of the siphons. Middle: Mussels, sessile bivalves, have a reduced forward muscle, the scallops have lost it altogether. Below: Specialized burrowing forms.
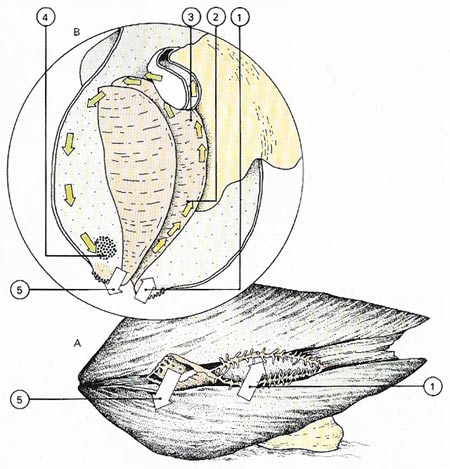
Figure 8. Bivalves are filter feeders (A); they extract food from water drawn into an entry siphon (1). By viewing a clam with its shell removed (B) it can be seen that the water moves across mucus-coated gills (2). The food particles in the water are trapped by the mucus and swept by ciliary action (3) towards the mouth. The filtered water and rejected food (4) move out through the discharge siphon (5).
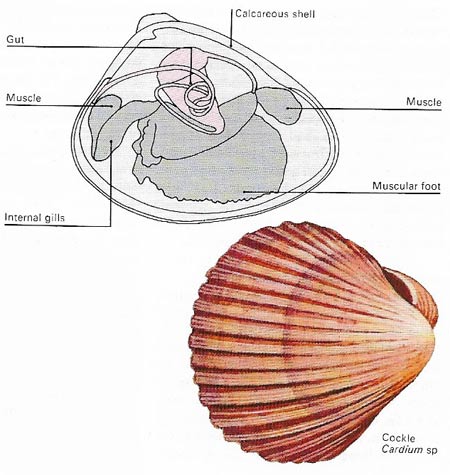
Figure 9. Bivalves have calcareous shells, the two halves of which are hinged, and can be closed by muscular action. The gills are used for both respiration and feeding – food trapped by the gills is passed to the mouth and gut by hair-like cilia. The muscular foot is used during burrowing.
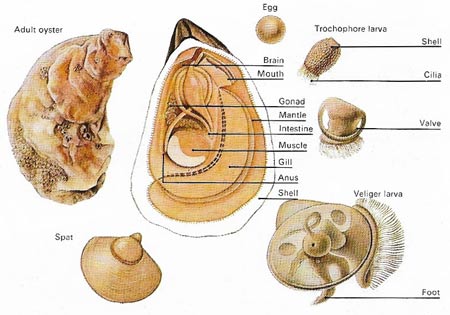
Figure 10. In oysters, as in most bivalves, the sexes are separate. Eggs and sperm are shed into the water and after fertilization the embryo develops into a free-swimming trochophore larva with vestiges of a shell and a bundle of cilia for propulsion and feeding. As the shell enlarges so the second or veliger larval form develops. This has a well-muscled foot, a shell and internal organs. The spat or young oyster remains free-swimming for some two weeks before settling down and attaching itself to a suitable surface. Some bivalves go through a brief parasitic phase that aids dispersal. The larva of these species – a glochidium – must attach itself to a fish for development to take place. Afterwards the young bivalve abandons its host to take up an independent existence.
A bivalve is any animal of the class Bivalvia (also called Pelecypoda) – one of the three main groups of molluscs. Bivalves are laterally compressed and possess a shell composed of two valves that hinge along their back, or dorsal, margin and enclose the body. Approximately 30,000 species are known, including such familiar examples as clams, oysters, and mussels.
Introduction
At a first inspection, the differences between a cockle and a snail seem to outnumber the likenesses. In fact, there hardly seems to be any resemblance between the two animals at all. Yet the snail (a gastropod) and the cockle (a bivalve or lamellibranch) are structurally very similar. They both belong to the Mollusca – one of the great divisions of the animal kingdom. Both have a hard protective shell, a mantle (a tent-like flap of tissue surrounding the body), a cavity within the mantle containing gills, and a powerful muscular foot.
The ancient molluscs, ancestors of bivalves, gastropods and the other molluscan animals (cephalopods, scaphopods and amphineura) most probably crawled about in shallow water feeding off minute organisms which they lapped up using a horny, rasping tongue or radula. Water percolating through the mantle cavity, washed against the gills providing oxygen.
The gastropods have remained closer to this original way of life. They still move about looking for food, and possess a distinct head armed with a horny radula. But the bivalves have moved a long way from the ancestral mode of living. They have become filter feeders. Exclusively aquatic creatures, they waft fine food particles into their mouths using tiny hairs or cilia usually arranged on the gills.
The gills, increasing in importance, have also increased in size. The mantle cavity which houses them has become elongated, running down either side of the animal, often for almost the entire length. Bivalves are therefore usually long and compressed from side to side.
The mantle itself has become divided into two lobes. Each lobe secretes a segment of shell, so the final structure is made of two valves – not, a single structure.
But the mantle does remain slightly connected at the top (dorsal) surface. Here a ligament – an elastic strip made of organic material – is secreted, either on the inside or the outside of the hinge. Two muscles run between the valves, a forward anterior adductor and a backward posterior adductor. When the muscles contract, the valves close together but when the muscles relax the elastic ligament at the hinge causes the shell to grape.
Bivalves, as filter feeders, do not really have a distinct head like snails. The mouth is at the front (anterior) end of the animal surrounded by two long bilobed palps (labial palps) but no sense organs are concentrated here and there is no rasping radula.
Structure of bivalves
The bulk of the bivalve body lies toward the hinge and is covered by a structure called the mantle. This grows outward on either side of the body and forms a lining to the inside of both shell halves. Indeed, it is the tissue lining or epithelium of the mantle that is responsible for secreting the shell. A muscular foot – flattened from side to side, extensible, and often used in burrowing – projects from the body into the cavity separating the halves of the mantle.
If one shell and half the mantle of a bivalve are removed the most obvious structures exposed are the gills which, besides serving as organs of respiration, are used in feeding. Bivalves are filter feeders. Tubular extensions of the mantle form a pair of siphons, one for the entry of a current of water, the other for its exit. This current is maintained by the beating action of hair-like cilia with which the gills are liberally covered. Water is drawn in through tiny pores in the gills and eventually ejected by way of the discharge siphon. Particles of food are strained out and trapped in a layer of sticky mucus, a substance similar in consistency to human saliva, that covers the gill surfaces.
From there, trapped food passes to the edge of the gills farthest away from the hinge, and larger particles, such as silt, fall off into the mantle cavity. The remainder, still entangled in a string of mucus, moves forward to the palps surrounding the mouth. Further sorting then takes place and selected particles enter the mouth and pass into the stomach for digestion.
Digestion and reproduction
A gland surrounding the stomach is the chief organ of digestion in bivalves, but projecting into the stomach, from an intestinal pouch at one end of it, is the crystalline style, a gelatinous rod containing carbohydrate-splitting enzymes. Cilia in the pouch make this rod rotate continually and its free end rubs against an area of the stomach known as the gastric shield. As the crystalline style is worn away so the enzymes it contains are released into the stomach contents. Digestion is completed in the intestine, which follows a tortuous course to the anus. This opens into the mantle cavity near the discharge siphon, thus ensuring the removal of feces in the outgoing water.
Paired excretory organs lie on either side of the heart and a "kidney" in each extracts waste products and discharges them, via a bladder, into the gill passages and thence to the exterior. Bivalves have a three-chambered heart whose single muscular ventricle pumps blood into arteries that run to the foot, mantle, and the body organs. Many of the tissues of the body are permeated not by capillaries, as in higher animals, but by blood spaces or sinuses. Sinuses in the foot enabling sand-burrowing – the foot swells and elongates as extra blood is pumped into them. Blood then returns via a series of veins to the paired atrial chambers of the heart.
Bivalves have simple reproductive systems with paired gonads and no glands. The sexes are separated and most allow the spermatozoa and ova to escape into the water where, in marine species, they form part of the plankton. Some freshwater bivalves brood their young, while others release a larval stage called a glochidium that passes through a brief parasitic phase before assuming an independent adult existence.
Gill structure
In snails the gills are simple, consisting of a stalk giving off plume-like filaments – rather like feathers. But in the bivalves the filaments become joined up forming a solid or plate-like structure (lammellibranchiata in fact means, plate-gilled).
In a very few simple bivalves (Protobranches) the gills remain confined to a cavity at the rear of the body and the filaments are short. In more advanced forms each filament becomes elongated and turns upwards. The gills are also extended so that they run practically the length of the body. Water is drawn into the shell at the rear (posterior) end where the mantle is often extended backwards to form two siphons. The lower siphon takes the water in. The water not only provides oxygen. Particles of food in the water are swept forward and downwards moving along the bottom of the gill filaments by rapidly beating of tracts of cilia. During the passage forward the particles become entangled in mucus.
From the gills food particles are swept onto the labial palps about the mouth. The labial palps are also ciliated. Material may be swept into the mouth or alternately rejected. passed back by cilia action along the base of the gills, and washed out through the upper of the two siphons. Sorting at the labial palps selects the finest particles, not necessarily the most nutritious. Large, coarse particles are rejected.
Methods of movement
Sedentary bivalves, such as mussels, spend their lives attached to a form base such as a rock by a bundle of threads called the byssus. These threads are secreted by a gland in the foot and anchor their owner with surprising strength – a necessity if the animal is to avoid being swept away by the force of the waves. A more common mode of life, found in bivalves such as the cockles and razor shells, is sand-burrowing by means of a mobile foot. The depth of burrowing varies among species but the deeper penetrating forms often have long siphons for obtaining surface water for respiration and feeding.
In contrast to the sand-burrowing species, the shipworm burrows into wood by back and forth rotation of its shells – it literally drills its way forward. Some piddocks can even burrow into rock. The piddock must periodically cease borrowing and project the edge of the mantle on to the shell surface to deposit a fresh layer of calcium carbonate.
In most bivalves the head is reduced or absent and sensory organs are generally insignificant. But the animals do react to waterborne chemicals and to light. Some of the razor shells in particular are extremely sensitive to vibration and will vanish rapidly down their burrows if they are disturbed.
Three groups of bivalves
Bivalve molluscs, like snails retain a muscular foot. The foot lies between the two sets of gills on the underneath (ventral) side of the body. Its shape and position is altered by muscular contraction and the structure can be protruded through the gape of the shell.
In one group of bivalves – the so-called Normal bivalves – the foot is well developed and the creatures use it to move about at the surface (e.g. cockles) or in some instances to burrow (e.g. razor shells). The giant clams of the Pacific and Indian Oceans belong to this group. They may grow to be more than a yard in length. Food is not only obtained by filtering. Masses of small green unicellular algae grow in the tissues of the clam and sunlight if focused upon them by numerous lenses. Some of the algae are periodically digested.
The sessile group of bivalves have, in general, lost their mobility in the adult animal. Instead they have become firmly attached to the surface. Modifications have accompanied this stationary mode of life. The ventral foot has tended to move more and more towards the front of the animal. As a direct result the anterior adductor muscle has become small or is lost; usually each valve becomes highly asymmetrical in shape.
A byssus nearly always appears at some stage in the life history. A byssus is a mass of sticky, diverging threads arising from a pit at the back of the foot. In the mussel the byssus protrudes through the gape in the valves. The pearl oyster has come to lie on its right valve and the byssus emerges through a notch.
The common oysters have lost the foot and byssus altogether; the right valve becomes cemented to the ground.
Pecten, the scallop, is structurally a sessile bivalve. The front adductor muscle has been lost by the forward migration of the foot. But the remaining muscle has become enlarged. By rapid contraction the valves are clapped together and water is violently discharged. This is an efficient mechanism for swimming and Pecten has given up its sessile habit.
The last division of the bivalves is the deep-burrowers. Unlike the burrowers of the Normal group, the deep burrowers have become extremely modified to their mode of life. The valves are only weakly hinged; often they have completely lost their ligament and the valves permanently gape. The siphons are enormous, often dwarfing the rest of the body.

